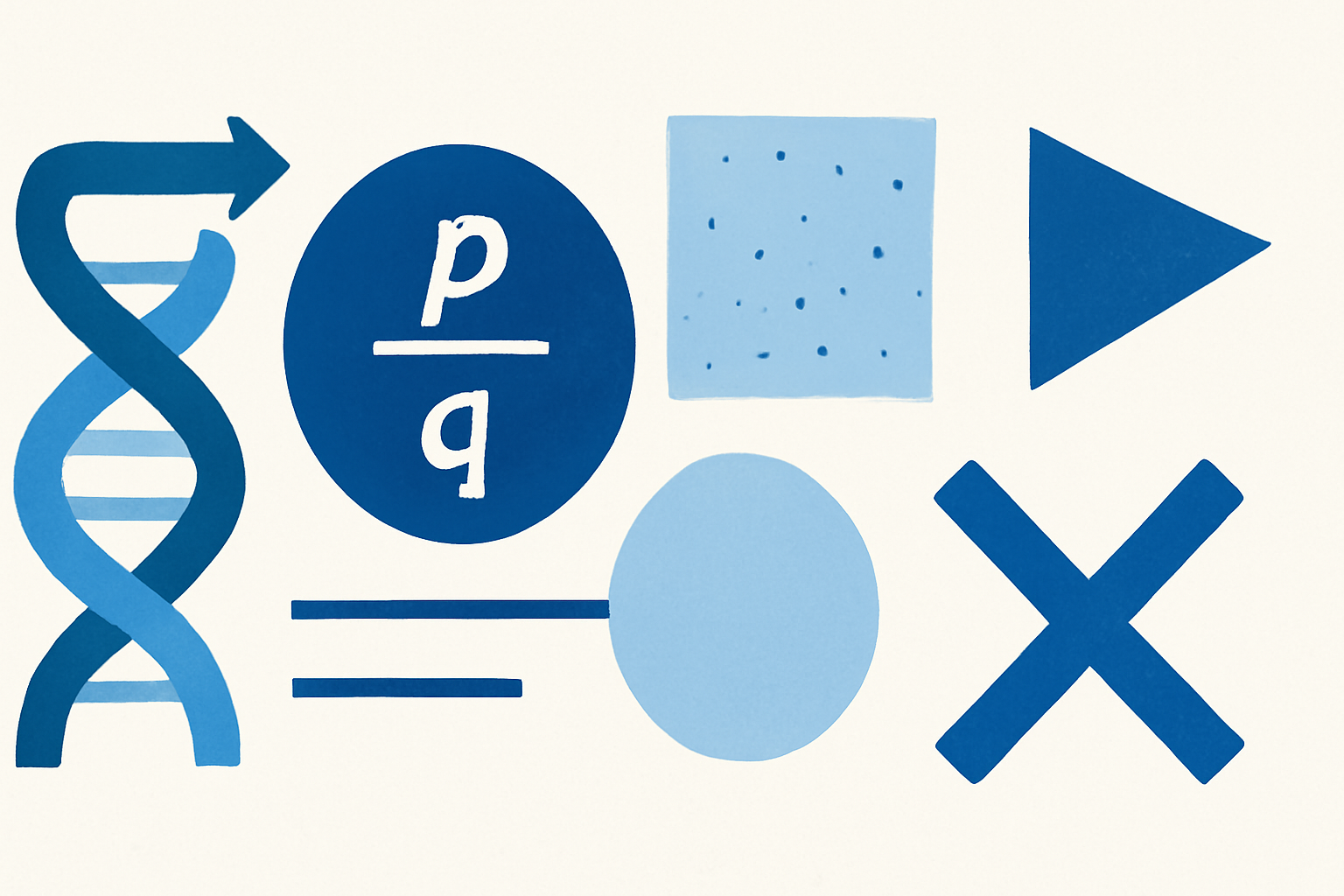
What the Hardy-Weinberg Equilibrium Actually Assumes
Population genetics rests on a deceptively simple equation, one that describes what a gene pool looks like when nothing interesting is happening. Understanding why that baseline matters—and what disturbs it—is central to understanding how evolution actually works.
A law for an imaginary population
In 1908, mathematician G. H. Hardy and physician Wilhelm Weinberg independently derived the same result: in a large, randomly mating population free from mutation, migration, and natural selection, allele frequencies stay constant from one generation to the next. The resulting Hardy-Weinberg equilibrium (HWE) gives exact predictions for how those frequencies translate into genotype frequencies. If an allele occurs at frequency p, and its counterpart at frequency q (where p + q = 1), then the expected genotype frequencies are p², 2pq, and q² for the three possible pairings.
The equation is algebraically trivial. Its importance is not algebraic. HWE defines a null model—what the gene pool would look like if evolution were not occurring. Any departure from predicted frequencies signals that at least one of the underlying assumptions is being violated. The law is useful precisely because real populations always violate it in one way or another.
What the assumptions actually require
HWE depends on five conditions, each worth examining on its own terms.
Random mating means every individual is equally likely to pair with every other. Humans do not mate randomly: we preferentially choose partners by geography, culture, language, and phenotype. This is called assortative mating, and it systematically alters genotype frequencies without changing allele frequencies—a subtle but important distinction.
No mutation is obviously never true. Mutation rates are low enough that their effect over a few generations is negligible, but over geological time mutation is the ultimate source of all genetic variation. Treating it as absent is a valid short-term approximation, not a claim about reality.
No migration means no gene flow in or out of the population. In practice, populations are rarely fully isolated. Gene flow tends to homogenize allele frequencies across populations, working against local adaptation. It is also one reason why clear genetic boundaries between human populations are far harder to draw than folk taxonomy suggests.
No genetic drift requires an infinitely large population—a condition no real population satisfies. In small populations, allele frequencies fluctuate randomly from generation to generation simply because reproduction is a sampling process. This is genetic drift, and its effects can be dramatic: rare alleles can be lost entirely by chance, or fixed in the population with no help from natural selection. The founder effect and population bottlenecks are specific instances of drift that have shaped the genetic diversity of nearly every species studied, including our own.
No natural selection means all genotypes must survive and reproduce equally. Selection is the mechanism Darwin identified, and it is omnipresent in living systems. When selection acts on a locus, genotype frequencies shift in a predictable direction—carriers of favoured alleles leave more offspring.
How HWE is used in practice
Because HWE defines a baseline, deviations from it are diagnostic. Genome-wide association studies (GWAS) routinely test each genetic marker for HWE before analysis. A marker that deviates significantly is often flagged as a potential genotyping error—if a heterozygote excess or deficit appears across thousands of samples, the most likely culprit is a technical artefact rather than genuine biology. HWE thus functions as a quality-control filter in large-scale genomic research.
In forensic genetics, HWE underpins the statistical models used to calculate the probability that a DNA profile would appear by chance in a population. If those models assumed non-equilibrium conditions, the reported match probabilities—often presented as astronomical odds—would need corresponding adjustment.
Conservation biology uses HWE violations in the opposite direction: as genuine biological signals. A captive population showing excess homozygosity may be experiencing inbreeding or drift, both of which erode adaptive potential. Managers use HWE analysis to decide whether and how to introduce individuals from other populations.
Why the null model is scientifically honest
It might seem strange that a law built on conditions that never hold is treated as foundational. But this is standard scientific practice, and it is intellectually honest rather than naive. A null model does not claim to describe the world; it describes what the world would look like in the absence of specific causal forces. Every confirmed departure from HWE is an invitation to ask which assumption is failing, and by how much.
This is exactly what good science looks like: building a precise, testable baseline and then measuring the gap between prediction and observation. The gap is where the biology lives. Hardy-Weinberg equilibrium is never actually achieved—and that is precisely what makes it worth calculating.


